
This page features selected student works from the studio. Documentation for each of the three projects includes a variety of precedent background, drawings, renderings, and photographs of models. Processing sketches, which develop dynamically and are interactive to mouse-overs and mouse-clicks, are also included for Project 1 as part of the students’ productions. Video demos of the sketches are provided in addition to the executable and code.
| TOPICS / PRECEDENTS | PROJECT COMPONENTS | |
|---|---|---|
| Project 1: “The Drawing Machine” | ||

Processing sketch with logic simulating the transformations of mesenchymal cells. |
EMT Variations in Cancer (Courtesy of MIT Student. Used with permission.) |
Video Demonstration of Project 1: EMT Variations in Cancer Executable for Windows (ZIP - 1.3MB) |
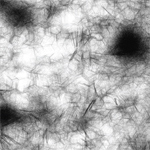
Processing sketch with logic simulating clusters of hexagonal and branching networks. |
Viral Capsid Self Assembly: Hierarchal Pair Construction (Courtesy of Phillip Hu. Used with permission.) |
Executable for Windows 32-bit (ZIP - 6.3MB) Executable for Windows 64-bit (ZIP - 6.3MB) Executable for Mac OS X (ZIP - 6.5MB) |

Processing sketch with logic simulating the Belousov-Zhabotinsky Reaction through drawn lines and network points. |
The Belousov-Zhabotinsky Reaction (Courtesy of Juanita Ballesteros. Used with permission.) |
Video Demonstration of Project 1: Belousov-Zhabotinsky Reaction |

Processing sketch with logic simulating the process of chitin synthesis. |
Chitin and Butterflies (Courtesy of Lina Kara’in. Used with permission.) |
Video Demonstration of Project 1: Chitin and Butterflies Executable for Windows 32-bit (ZIP - 2.4MB) Executable for Windows 64-bit (ZIP - 2.4MB) Executable for Mac OS X (ZIP - 2.8MB) |
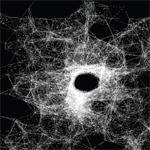
Processing sketch with logic simulating how a spider web reacts to a point of stress. |
Spider Web Deformation (Courtesy of Tiandra Ray. Used with permission.) |
Video Demonstration of Project 1: Spider Web Deformation Executable for Windows (ZIP - 1.3MB) |
| Project 2: “The Making Machine” | ||

Model of cell transformations as oxygen concentrations vary in the z-direction. |
EMT Variations in Cancer (Courtesy of MIT Student. Used with permission.) |
Documentation (PDF - 1.4MB) |

Model of branch growth shown through cylindrical branches and nodes. |
Branching Out (Courtesy of Marianna Gonzalez. Used with permission.) |
Documentation (PDF - 3.0MB) |
| Project 3: “The Architecture Machine” | ||

Final model depicting spatial relationships between labs, CO2 collection chambers, and vortex rings. |
CO2 Accumulation & Conversion (Courtesy of Anonymous Student. Used with permission.) |
Documentation (PDF - 9.0MB) |
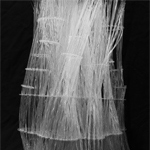
Final model of branch growth using plexiglass plates and rods. |
Branching Out (Courtesy of Marianna Gonzalez. Used with permission.) |
Documentation (PDF - 7.5MB) |
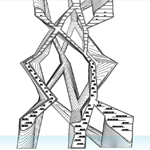
Section drawing of the branched spaces with partial submersion in water. |
Chitin and Butterflies (Courtesy of Lina Kara’in. Used with permission.) |
Documentation (PDF - 1.9MB) |
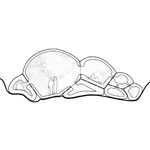
Section drawing of inhabitable spaces formed by packed volumes and their intersections. |
Packing, Volume, & Density (Courtesy of Christiana Rosales. Used with permission.) |
Documentation (PDF - 41.2MB) |





